CBP has two critical mechanisms by which it is able to regulate gene expression: as an acetyltransferase, and as a protein scaffold
This gene is ubiquitously expressed and is involved in the transcriptional coactivation of many different transcription factors. CBP has two critical mechanisms by which it is able to regulate gene expression: as an acetyltransferase, and as a protein scaffold that helps recruit and construct the complexes that are necessary for transcription or chromatin remodeling. Phosphorylation of CBP increases its acetyltransferase activity, a process hypothesized to be regulated in a cell cycle dependent manner. Recent results suggest that novel CBP-mediated post-translational N-glycosylation activity alters the conformation of CBP-interacting proteins, leading to regulation of gene expression, cell growth and differentiation.
Dutta R, Tiu B, Sakamoto KM (September 2016). “CBP/p300 acetyltransferase activity in hematologic malignancies”. Molecular Genetics and Metabolism. 119 (1–2): 37–43. doi:10.1016/j.ymgme.2016.06.013. PMID27380996.
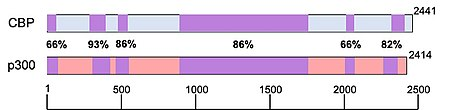
Often in scientific papers (especially in less recent ones), CBP and p300 are used interchangeably as CBP/p300. This is a reasonable amalgamation given the sequence homology, structural similarity and binding behaviors of these two proteins. However, ongoing research shows that CBP and p300 maintain distinct biological functions and should therefore not be conflated.
It has been reported that despite having common histone substrates, when there are lower amounts of histone or acetyl-CoA available, CBP and p300 have different, preferred acetyltransferase substrates. In an experiment performed involving the Kaposi sarcoma-associated herpesvirus, the pathological protein (vIRF), was shown to be upregulated by CBP and repressed by p300. p300 homozygous knockout studies in mice were embryonic lethal, with improper neurulation and poor heart development occurring during their limited survival. Additionally, the fibroblasts isolated from these mice were unable to properly proliferate and were lacking the retinoic acid receptor. Transgenically altered mice homozygous for mutated copies of CBP (missing the KAT domain), were also embryonic lethal, however instead, these mice had poor vascular angiogenesis and abnormal hematopoiesis characterized by the absence of progenitor cell proliferation and an altered hematopoietic microenvironment. The fact that both CBP and p300 homozygous knockouts were embryonic lethal suggest that these factors play a critical role in embryogenesis. The differential phenotypes of these embryos also indicate that CBP and p300 regulate different aspects of embryological development.
Dutta R, Tiu B, Sakamoto KM (September 2016). “CBP/p300 acetyltransferase activity in hematologic malignancies”. Molecular Genetics and Metabolism. 119 (1–2): 37–43. doi:10.1016/j.ymgme.2016.06.013. PMID27380996.
Role in cell cycle regulation
Studies performed in the late 1990s demonstrated that peak CBP acetyltransferase activity occurs at the transition between the G1/S phase of the cell cycle checkpoint. Given the concentrations of CDK2 present in the cell at this stage of the cell cycle, it was hypothesized that CDK2 may be a key regulator of these post-translational modifications. It turned out that administration of cyclin E/CDK2 inhibitors did in fact inhibit the enzymatic activity of CBP’s KAT domain. Other proteins that have been shown to phosphorylate CBP include MAP Kinase, PKA and CAMK4. Ser-133 has been shown to be an important residue that is phosphorylated by PKA to initiate CBP transcriptional activity.
Ait-Si-Ali S, Ramirez S, Barre FX, Dkhissi F, Magnaghi-Jaulin L, Girault JA, et al. (November 1998). “Histone acetyltransferase activity of CBP is controlled by cycle-dependent kinases and oncoprotein E1A”. Nature. 396 (6707): 184–186. Bibcode:1998Natur.396..184A. doi:10.1038/24190. PMID9823900. S2CID4340781.
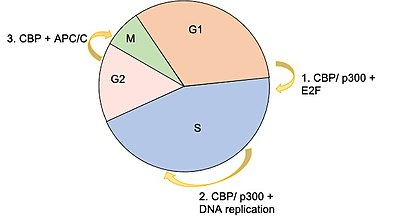
The E2F family of transcription factors are critical for taking a cell from G1 to S phase of the cell cycle. They bind to a sequence consensus in the promoter region of genes involved in DNA replication. CBP (and p300) have been shown to interact with E2F proteins as both a coactivator, and as an acetyltransferase, the latter of which causes increased E2F DNA binding affinity. A knockout study that was published in 2000, which used a microinjection of an antibody against CBP/p300 significantly diminished the number of cells able to progress into S phase, further supporting the idea that CBP is essential in the transcription of factors necessary G1/S phase transition.
Chan HM, La Thangue NB (July 2001). “p300/CBP proteins: HATs for transcriptional bridges and scaffolds”. Journal of Cell Science. 114 (Pt 13): 2363–2373. doi:10.1242/jcs.114.13.2363. PMID11559745.
CBP is also believed to facilitate the process of DNA replication during S phase by acetylating histones around origins of replication. Acetylation of histones, specifically lysine residues in the histones, weakens the electric charge interaction between the histone and DNA, causing this area to become more open and accessible for the machinery required for DNA replication. Two histone acetylation markers that have been associated with active regions include histone 3 lysine 18 acetylation (H3K18ac) and histone 3 lysine 27 acetylation (H3K27ac). It has also been shown that CBP acetylates two endonucleases (FEN1, DNA2) that are involved in processing Okazaki fragments.
Another key component of the cell cycle that is regulated by CBP is the anaphase promoting complex/cyclosome (APC/C). This complex consists of numerous subunits that are grouped into two subdomains, the “Arc Lamp” and the “Platform,” and functions as an E3 ubiquitin ligase that targets components related to the cell cycle such as cyclin B, securin, and PLK1 for proteasome degradation. Two subunits of the APC/C have been shown to directly interact with CBP: AP5, which is located in the Platform subdomain, and AP7, located in the Arc Lamp subdomain. Experiments performed using RNAi to completely suppress CBP and p300 showed significant increases in protein concentrations for those normally targeted by the APC/C, and caused a number of cells to become arrested in the mitotic phase of the cell cycle.
Turnell AS, Stewart GS, Grand RJ, Rookes SM, Martin A, Yamano H, et al. (December 2005). “The APC/C and CBP/p300 cooperate to regulate transcription and cell-cycle progression”. Nature. 438 (7068): 690–695. Bibcode:2005Natur.438..690T. doi:10.1038/nature04151. PMID16319895. S2CID4432156.
Dutto I, Scalera C, Prosperi E (April 2018). “CREBBP and p300 lysine acetyl transferases in the DNA damage response”. Cellular and Molecular Life Sciences. 75 (8): 1325–1338. doi:10.1007/s00018-017-2717-4. PMID29170789. S2CID3951961.
{kind=link}
Leave a Reply