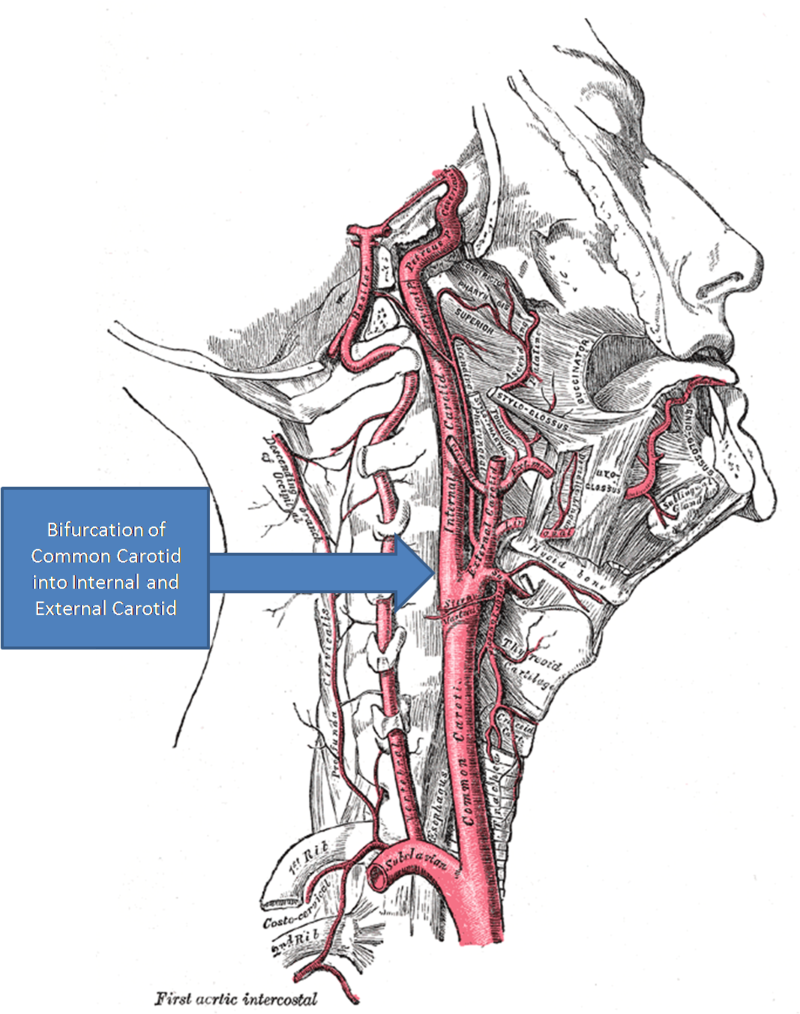
The carotid body is not labelled but is located within the wall of the internal carotid artery near the bifurcation of the common carotid artery. Glomus type I cells are the chemoreceptor cells found in the carotid body.
Glomus cells are the cell type mainly located in the carotid bodies and aortic bodies. Glomus type I cells are peripheral chemoreceptors which sense the oxygen, carbon dioxide and pH levels of the blood. When there is a decrease in the blood’s pH, a decrease in oxygen (pO2), or an increase in carbon dioxide (pCO2), the carotid bodies and the aortic bodies signal the dorsal respiratory group in the medulla oblongata to increase the volume and rate of breathing.
Lahiri S, Semenza G, Prabhakar NR, eds. (2003). Oxygen sensing : responses and adaptation to Hypoxia. New York: Dekker. pp. 200, 232. ISBN978-0824709600.
The glomus cells have a high metabolic rate and good blood perfusion and thus are sensitive to changes in arterial blood gas tension. Glomus type II cells are sustentacular cells having a similar supportive function to glial cells.
Pearse AG, Polak JM, Rost FW, Fontaine J, Le Lièvre C, Le Douarin N (1973). “Demonstration of the neural crest origin of type I (APUD) cells in the avian carotid body, using a cytochemical marker system”. Histochemie. 34 (3): 191–203. doi:10.1007/bf00303435. PMID4693636. S2CID25437552.
Lawson, W (January 1980). “The neuroendocrine nature of the glomus cells: an experimental, ultrastructural, and histochemical tissue culture study”. The Laryngoscope. 90 (1): 120–44. doi:10.1288/00005537-198001000-00014. PMID6243386. S2CID13149248.
Eyzaguirre, C; Fidone, SJ (November 1980). “Transduction mechanisms in carotid body: glomus cells, putative neurotransmitters, and nerve endings”. The American Journal of Physiology. 239 (5): C135-52. doi:10.1152/ajpcell.1980.239.5.C135. PMID6108075.
CB glomus cells have also been found to detect hypoglycemia in both non-primate mammals and humans. O2 and low-glucose responses share a common final pathway involving membrane depolarization, extracellular calcium influx, increase in cytosolic calcium concentration, and neurotransmitter secretion, which stimulates afferent sensory fibers to evoke sympathoadrenal activation. On the other hand, hypoxia and low glucose induce separate signal transduction pathways. Unlike O2 sensing, the response of the CB to low glucose is not altered by rotenone, with the low glucose-activated background cationic current unaffected by hypoxia. Responses of the CB to hypoglycemia and hypoxia can be potentiated by each other. The counter-regulatory response to hypoglycemia by the CB is essential for the brain, an organ that is particularly sensitive to low glucose. CB glucose sensing could be altered in diabetic patients, particularly those under insulin treatment, as well as in other medical conditions such as sleep apnea or obstructive pulmonary diseases, where chronic hypoxemia presents with plastic modifications in CB structure and function.
Lin Gao, Glucose sensing by carotid body glomus cells: potential implications in disease. Front. Physiol., 15 October 2014 Sec. Integrative Physiology Volume 5 – 2014 | https://doi.org/10.3389/fphys.2014.00398
Glomus type I cells are chemoreceptors which monitor arterial blood for the partial pressure of oxygen (pO2), partial pressure of carbon dioxide (pCO2) and pH.
Carroll, JL; Kim, I (15 November 2005). “Postnatal development of carotid body glomus cell O2 sensitivity”. Respiratory Physiology & Neurobiology. 149 (1–3): 201–15. doi:10.1016/j.resp.2005.04.009. PMID15886071. S2CID25277654.
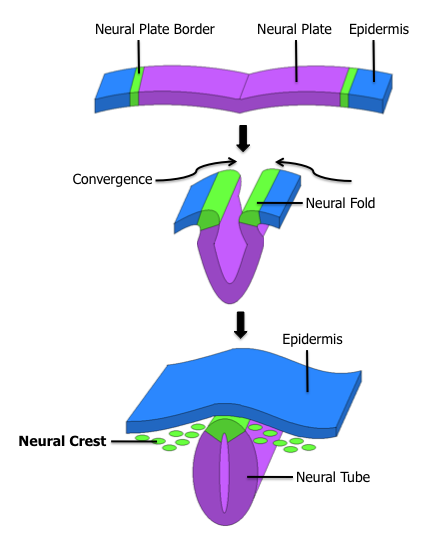
Development of the nervous system. The glomus type I cells of the carotid body are derived from the neural crest and can be seen in green.
Glomus type I cells are embryonically derived from the neural crest. In the carotid body the respiratory chemoreceptors need a period of time postnatally in order to reach functional maturity. This maturation period is known as resetting.
Pearse AG, Polak JM, Rost FW, Fontaine J, Le Lièvre C, Le Douarin N (1973). “Demonstration of the neural crest origin of type I (APUD) cells in the avian carotid body, using a cytochemical marker system”. Histochemie. 34 (3): 191–203. doi:10.1007/bf00303435. PMID4693636. S2CID25437552.
At birth the chemorecptors express a low sensitivity for lack of oxygen but this increases over the first few days or weeks of life. The mechanisms underlying the postnatal maturity of chemotransduction are obscure.
Carroll, JL; Kim, I (15 November 2005). “Postnatal development of carotid body glomus cell O2 sensitivity”. Respiratory Physiology & Neurobiology. 149 (1–3): 201–15. doi:10.1016/j.resp.2005.04.009. PMID15886071. S2CID25277654.
Clinical significance
Clusters of glomus cells, of which the carotid bodies and aortic bodies are the most important, are called non-chromaffin or parasympathetic paraganglia. They are also present along the vagus nerve, in the inner ears, in the lungs, and at other sites. Neoplasms of glomus cells are known as paraganglioma, among other names, they are generally non-malignant.
Anne Marie McNicol (2010). “Chapter 12: Adrenal medulla and paraganglia”. Endocrine Pathology: Differential Diagnosis and Molecular Advance (Springer ed.). p. 281.
Lahiri S, Semenza G, Prabhakar NR, eds. (2003). Oxygen sensing : responses and adaptation to Hypoxia. New York: Dekker. pp. 200, 232. ISBN978-0824709600.
Pearse AG, Polak JM, Rost FW, Fontaine J, Le Lièvre C, Le Douarin N (1973). “Demonstration of the neural crest origin of type I (APUD) cells in the avian carotid body, using a cytochemical marker system”. Histochemie. 34 (3): 191–203. doi:10.1007/bf00303435. PMID4693636. S2CID25437552.
Lawson, W (January 1980). “The neuroendocrine nature of the glomus cells: an experimental, ultrastructural, and histochemical tissue culture study”. The Laryngoscope. 90 (1): 120–44. doi:10.1288/00005537-198001000-00014. PMID6243386. S2CID13149248.
Eyzaguirre, C; Fidone, SJ (November 1980). “Transduction mechanisms in carotid body: glomus cells, putative neurotransmitters, and nerve endings”. The American Journal of Physiology. 239 (5): C135-52. doi:10.1152/ajpcell.1980.239.5.C135. PMID6108075.
Carroll, JL; Kim, I (15 November 2005). “Postnatal development of carotid body glomus cell O2 sensitivity”. Respiratory Physiology & Neurobiology. 149 (1–3): 201–15. doi:10.1016/j.resp.2005.04.009. PMID15886071. S2CID25277654.
Anne Marie McNicol (2010). “Chapter 12: Adrenal medulla and paraganglia”. Endocrine Pathology: Differential Diagnosis and Molecular Advance (Springer ed.). p. 281.
Leave a Reply