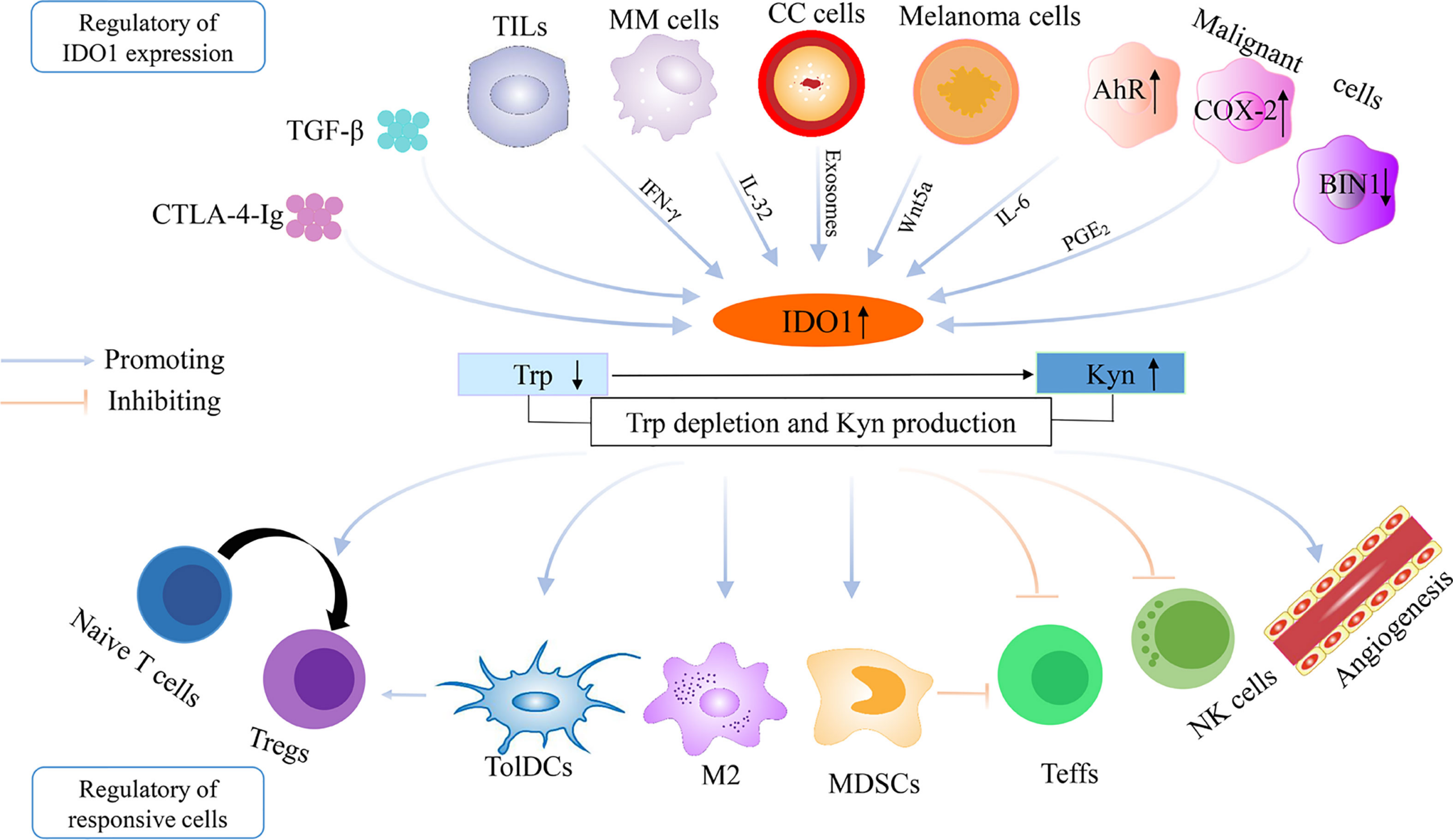
IDO is an important part of the immune system and plays a part in natural defense against various pathogens. It is produced by the cells in response to inflammation and has an immunosuppressive function because of its ability to limit T-cell function and engage mechanisms of immune tolerance.
Emerging evidence suggests that IDO becomes activated during tumor development, helping malignant cells escape eradication by the immune system. Expression of IDO has been described in a number of types of cancer, such as acute myeloid leukemia, ovarian cancer or colorectal cancer. IDO is part of the malignant transformation process and plays a key role in suppressing the anti-tumor immune response in the body, so inhibiting it could increase the effect of chemotherapy as well as other immunotherapeutic protocols.
Uyttenhove C, Pilotte L, Théate I, Stroobant V, Colau D, Parmentier N, et al. (October 2003). “Evidence for a tumoral immune resistance mechanism based on tryptophan degradation by indoleamine 2,3-dioxygenase”. Nature Medicine. 9 (10): 1269–74. doi:10.1038/nm934. PMID14502282. S2CID10618102.
Physiological function
Indoleamine 2,3-dioxygenase is the first and rate-limiting enzyme of tryptophancatabolism through the kynurenine pathway.
IDO is an important molecule in the mechanisms of tolerance and its physiological functions include the suppression of potentially dangerous inflammatory processes in the body.
Romani L, Fallarino F, De Luca A, Montagnoli C, D’Angelo C, Zelante T, et al. (January 2008). “Defective tryptophan catabolism underlies inflammation in mouse chronic granulomatous disease”. Nature. 451 (7175): 211–5. Bibcode:2008Natur.451..211R. doi:10.1038/nature06471. PMID18185592. S2CID4391121.
IDO also plays a role in natural defense against microorganisms. Expression of IDO is induced by interferon-gamma, which explains why the expression increases during inflammatory diseases or even during tumorigenesis.
Since tryptophan is essential for the survival of pathogens, the activity of enzyme IDO destroys them. Microorganisms susceptible to tryptophan deficiency include bacteria of genus Streptococcus or viruses such as herpes simplex or measles.
One of the organs with high IDO expression is the placenta. In the 1990s, the immunosuppressive function of this enzyme was first described in mice due to the study of placental tryptophan metabolism. Thus, mammalian placenta, due to intensive tryptophan catabolism has the ability to suppress T cell activity, thereby contributing to its position of immunologically privileged tissue.
IDO is known to suppress T and NK cells (aka natural killer cells or large granular lymphocytes (LGL), a type of cytotoxic lymphocyte critical to the innate immune system that belong to the rapidly expanding family of known innate lymphoid cells (ILC) and represent 5–20% of all circulating lymphocytes in humans.), generate Tregs and myeloid-derived suppressor cells, and also supports angiogenesis.
Note: The regulatory T cells (Tregs or Treg cells), formerly known as suppressor T cells, are a subpopulation of T cells that modulate the immune system, maintain tolerance to self-antigens, and prevent autoimmune disease. Treg cells are immunosuppressive and generally suppress or downregulate induction and proliferation of effector T cells. Treg cells express the biomarkers CD4, FOXP3, and CD25 and are thought to be derived from the same lineage as naïve CD4+ cells. Because effector T cells also express CD4 and CD25, Treg cells are very difficult to effectively discern from effector CD4+, making them difficult to study. Research has found that the cytokine transforming growth factor beta (TGF-β) is essential for Treg cells to differentiate from naïve CD4+ cells and is important in maintaining Treg cell homeostasis.The immune system must be able to discriminate between self and non-self. When self/non-self discrimination fails, the immune system destroys cells and tissues of the body and as a result causes autoimmune diseases. Regulatory T cells actively suppress activation of the immune system and prevent pathological self-reactivity, i.e. autoimmune disease. The critical role regulatory T cells play within the immune system is evidenced by the severe autoimmune syndrome that results from a genetic deficiency in regulatory T cells (IPEX syndrome – Immune dysregulation, Polyendocrinopathy, Enteropathy X-linked (IPEX) syndrome).
Bettelli E, Carrier Y, Gao W, Korn T, Strom TB, Oukka M, et al. (May 2006). “Reciprocal developmental pathways for the generation of pathogenic effector TH17 and regulatory T cells”. Nature. 441 (7090): 235–8. Bibcode:2006Natur.441..235B. doi:10.1038/nature04753. PMID16648838. S2CID4391497.
Myeloid-derived suppressor cells (MDSC) are a heterogeneous group of immune cells from the myeloid lineage (a family of cells that originate from bone marrow stem cells). MDSCs expand under pathologic conditions such as chronic infection and cancer, as a result of altered haematopoiesis. MDSCs differ from other myeloid cell types in that they have immunosuppressive activities, as opposed to immune-stimulatory properties. Similar to other myeloid cells, MDSCs interact with immune cell types such as T cells, dendritic cells, macrophages and natural killer cells to regulate their functions. MDSCs are formed from bone marrow precursors when myelopoietic processes are interrupted, caused by several illnesses. Cancer patients’ growing tumors produce cytokines and other substances that affect MDSC development. Tumor cell lines overexpress colony-stimulating factors (G-CSF and GM-CSF) and IL6, which promote development of MDSCs that have immune suppressive function in vivo. Other cytokines, including IL10, IL1, VEGF, and PGE2 have been associated with the formation and regulation of MDSCs. GM-CSF promotes synthesis of MDSCs from bone marrow, and the transcription factor c/EBP regulates development of MDSCs in bone marrow and in tumors. STAT3 also promotes development of MDSCs, whereas IRF8 could counteract MDSC-inducing signals. MDSCs migrate as immature cells from the bone marrow to peripheral tissues (or tumors), where they differentiate into mature macrophages, dendritic cells, and neutrophils without suppressive phenotypes under homeostatic conditions, but become polarized when exposed to pro-inflammatory compounds, chemokines, and cytokines. In the tumor microenvironment, they suppress the anti-tumor immune response. The presence of MDSCs has been associated with progression of colon cancer, tumor angiogenesis, and metastases. In addition to producing NO and ROS, MDSCs secrete immune-regulatory cytokines such as TNF, TGFB, and IL10. There are subpopulations of MDSC that have some common suppressive characteristics but also have their own unique features; different subpopulations can be found in different areas of the same tissue or tumor. Tumor-infiltrating MDSCs develop in response to environmental factors, upregulating CD38 (which removes NAD from the environment and is necessary for mitochondrial biosynthesis), PDL-1 (an immune checkpoint protein) and LOX1 (promotes fatty acid consumption and fatty acid oxidation). Tumor-infiltrating MDSCs also secrete exosomes that can inhibit the anti-tumor immune response. MDSCs are immune suppressive and play a role in tumor maintenance and progression. MDSCs also obstruct therapies that seek to treat cancer through both immunotherapy and other non-immune means. MDSC activity was originally described as suppressors of T cells, in particular of CD8+ T-cell responses. The spectrum of action of MDSC activity also encompasses NK cells, dendritic cells and macrophages. Suppressor activity of MDSC is determined by their ability to inhibit the effector function of lymphocytes. Inhibition can be caused by different mechanisms. It is primarily attributed to the effects of the metabolism of L-arginine. Another important factor influencing the activity of MDSC is oppressive ROS. MDSCs can also play a positive regulatory role. It is stated that MMR vaccine stimulates MDSC populations in people taking the vaccine, inhibiting septic inflammation and mortality that is broadly applicable not only to measles, mumps, and rubella, but extends to covid-19 induced cytokine inflammation.[citation needed] This vaccination inducement appears to be neither permanent nor chronic.[clarification needed] Despite MDSC’s being immunosuppressive in certain instances, the MMR vaccine itself is immunostimulatory. The term myeloid-derived suppressor cell originated in a 2007 journal article published in Cancer Research by Gabrilovich et al. Publications in 2008 established that there are two subpopulations of MDSC: mononuclear MDSC (M-MDSC) and polymorphonuclear or granulocytic MDSC (PMN-MDSC). M-MDSC are similar to monocytes found in blood, while PMN-MDSC are physically akin to neutrophils.
Poschke I, Kiessling R (September 2012). “On the armament and appearances of human myeloid-derived suppressor cells”. Clinical Immunology. 144 (3): 250–268. doi:10.1016/j.clim.2012.06.003. PMID22858650.
Gabrilovich D (2013-01-01). “Abstract IA7: Regulation of myeloid-derived suppressor cells in tumor micro-environment”. Tumor Microenvironment. American Association for Cancer Research. 73: IA7. doi:10.1158/1538-7445.tumimm2012-ia7.
Prochetto E, Borgna E, Jiménez-Cortegana C, Sánchez-Margalet V, Cabrera G. Myeloid-derived suppressor cells and vaccination against pathogens. Front Cell Infect Microbiol. 2022 Sep 29;12:1003781. doi: 10.3389/fcimb.2022.1003781. PMID: 36250061; PMCID: PMC9557202.
Fernández A, Oliver L, Alvarez R, Fernández LE, Lee KP, Mesa C. Adjuvants and myeloid-derived suppressor cells: enemies or allies in therapeutic cancer vaccination. Hum Vaccin Immunother. 2014;10(11):3251-60. doi: 10.4161/hv.29847. PMID: 25483674; PMCID: PMC4514045.
These mechanisms are crucial in the process of carcinogenesis. IDO allows tumor cells to escape the immune system by two main mechanisms. The first mechanism is based on tryptophan depletion from the tumor microenvironment.
Expression of IDO is under physiological conditions regulated by the Bin1 gene, which can be damaged by tumor transformation.
Muller, Alexander J; DuHadaway, James B; Donover, P Scott; Sutanto-Ward, Erika; Prendergast, George C (2005-02-13). “Inhibition of indoleamine 2,3-dioxygenase, an immunoregulatory target of the cancer suppression gene Bin1, potentiates cancer chemotherapy”. Nature Medicine. 11 (3): 312–319. doi:10.1038/nm1196. ISSN1078-8956. PMID15711557. S2CID12338548.
Emerging clinical studies suggest that combination of IDO inhibitors with classical chemotherapy and radiotherapy could restore immune control and provide a therapeutic response to generally resistant tumors. Enzyme IDO used by tumors to escape immune surveillance is currently in focus of research and drug discovery efforts, as well as efforts to understand if it could be used as a biomarker for prognosis.
Jiang T, Sun Y, Yin Z, Feng S, Sun L, Li Z (2015). “Research progress of indoleamine 2,3-dioxygenase inhibitors”. Future Medicinal Chemistry. 7 (2): 185–201. doi:10.4155/fmc.14.151. PMID25686005.
Song X, Si Q, Qi R, Liu W, Li M, Guo M, Wei L, Yao Z. Indoleamine 2,3-Dioxygenase 1: A Promising Therapeutic Target in Malignant Tumor. Front Immunol. 2021 Dec 23;12:800630. doi: 10.3389/fimmu.2021.800630. PMID: 35003126; PMCID: PMC8733291.
Inhibitors
COX-2 inhibitors down-regulate indoleamine 2,3-dioxygenase, leading to a reduction in kynurenine levels as well as reducing proinflammatory cytokine activity.[citation needed]
1-Methyltryptophan is a racemic compound that weakly inhibits indoleamine dioxygenase, but is also a very slow substrate. The specific racemer 1-methyl-d-tryptophan (known as indoximod) is in clinical trials for various cancers.
Epacadostat (INCB24360), navoximod (GDC-0919), and linrodostat (BMS-986205) are potent inhibitors of the indoleamine 2,3-dioxygenase enzyme and are in clinical trials for various cancers.
Uyttenhove C, Pilotte L, Théate I, Stroobant V, Colau D, Parmentier N, et al. (October 2003). “Evidence for a tumoral immune resistance mechanism based on tryptophan degradation by indoleamine 2,3-dioxygenase”. Nature Medicine. 9 (10): 1269–74. doi:10.1038/nm934. PMID14502282. S2CID10618102.
Romani L, Fallarino F, De Luca A, Montagnoli C, D’Angelo C, Zelante T, et al. (January 2008). “Defective tryptophan catabolism underlies inflammation in mouse chronic granulomatous disease”. Nature. 451 (7175): 211–5. Bibcode:2008Natur.451..211R. doi:10.1038/nature06471. PMID18185592. S2CID4391121.
Muller, Alexander J; DuHadaway, James B; Donover, P Scott; Sutanto-Ward, Erika; Prendergast, George C (2005-02-13). “Inhibition of indoleamine 2,3-dioxygenase, an immunoregulatory target of the cancer suppression gene Bin1, potentiates cancer chemotherapy”. Nature Medicine. 11 (3): 312–319. doi:10.1038/nm1196. ISSN1078-8956. PMID15711557. S2CID12338548.
Jiang T, Sun Y, Yin Z, Feng S, Sun L, Li Z (2015). “Research progress of indoleamine 2,3-dioxygenase inhibitors”. Future Medicinal Chemistry. 7 (2): 185–201. doi:10.4155/fmc.14.151. PMID25686005.
{kind=link}
Leave a Reply