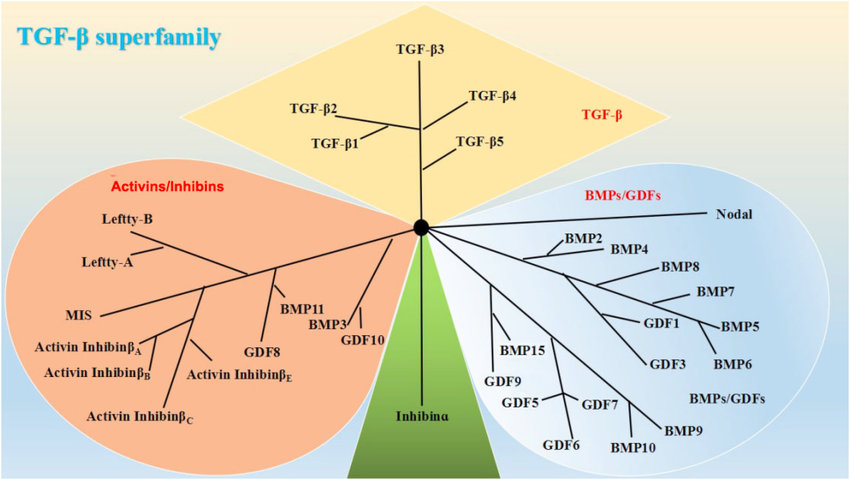
The TGF-β superfamily. Based on their structural features, the mammalian members of the TGF-β family are subdivided into (i) TGF-βs, (ii) activins/inhibins, and (iii) BMPs/growth and differentiation factors (GDFs). https://www.researchgate.net/publication/329884659_The_Role_of_TGF-b_and_Its_Receptors_in_Gastrointestinal_Cancers
The transforming growth factor beta (TGF-β) superfamily is a large group of structurally related cell regulatory proteins that was named after its first member, TGF-β1, originally described in 1983.
Unlike the Type I and II receptors which are kinases, TGFBR3 has a Zona pellucida-like domain. Its core domain binds TGF-beta family ligands and its heparan sulfate chains bind bFGF. It acts as a reservoir of ligand for TGF-beta receptors.
Andres JL, DeFalcis D, et al. (1992). “Binding of two growth factor families to separate domains of the proteoglycan betaglycan”. J. Biol. Chem. 267 (9): 5927–5930. PMID1556106.
Many proteins have since been described as members of the TGF-β superfamily in a variety of species, including invertebrates as well as vertebrates and categorized into 23 distinct gene types that fall into four major subfamilies.
Herpin A, Lelong C, Favrel P (May 2004). “Transforming growth factor-beta-related proteins: an ancestral and widespread superfamily of cytokines in metazoans”. Dev. Comp. Immunol. 28 (5): 461–85. doi:10.1016/j.dci.2003.09.007. PMID15062644.
Meng, Xiao-ming; Nikolic-Paterson, David J.; Lan, Hui Yao (June 2016). “TGF-β: the master regulator of fibrosis”. Nature Reviews Nephrology. 12 (6): 325–338. doi:10.1038/nrneph.2016.48. PMID27108839. S2CID25871413.
Activated TGF-β complexes with other factors to form a serine/threonine kinase complex that binds to TGF-β receptors. TGF-β receptors are composed of both type 1 and type 2 receptor subunits. After the binding of TGF-β, the type 2 receptor kinase phosphorylates and activates the type 1 receptor kinase that activates a signaling cascade. This leads to the activation of different downstream substrates and regulatory proteins, inducing transcription of different target genes that function in differentiation, chemotaxis, proliferation, and activation of many immune cells.
Nakao A, Afrakhte M, Morén A, Nakayama T, Christian JL, Heuchel R, et al. (October 1997). “Identification of Smad7, a TGFbeta-inducible antagonist of TGF-beta signalling”. Nature. 389 (6651): 631–5. Bibcode:1997Natur.389..631N. doi:10.1038/39369. PMID9335507. S2CID4311145.
TGF-β is secreted by many cell types, including macrophages, in a latent form in which it is complexed with two other polypeptides, latent TGF-beta binding protein (LTBP) and latency-associated peptide (LAP). Serum proteinases such as plasmin catalyze the release of active TGF-β from the complex. This often occurs on the surface of macrophages where the latent TGF-β complex is bound to CD36 via its ligand, thrombospondin-1 (TSP-1). Inflammatory stimuli that activate macrophages enhance the release of active TGF-β by promoting the activation of plasmin. Macrophages can also endocytose IgG-bound latent TGF-β complexes that are secreted by plasma cells and then release active TGF-β into the extracellular fluid. Among its key functions is regulation of inflammatory processes, particularly in the gut. TGF-β also plays a crucial role in stem cell differentiation as well as T-cell regulation and differentiation.
Because of its role in immune and stem cell regulation and differentiation, it is a highly researched cytokine in the fields of cancer, auto-immune diseases, and infectious disease.
The TGF-β superfamily includes endogenous growth inhibiting proteins; an increase in expression of TGF-β often correlates with the malignancy of many cancers and a defect in the cellular growth inhibition response to TGF-β. Its immunosuppressive functions then come to dominate, contributing to oncogenesis.
The dysregulation of its immunosuppressive functions is also implicated in the pathogenesis of autoimmune diseases, although their effect is mediated by the environment of other cytokines present.
Vaughn SP, Broussard S, Hall CR, Scott A, Blanton SH, Milunsky JM, Hecht JT (May 2000). “Confirmation of the mapping of the Camurati-Englemann locus to 19q13. 2 and refinement to a 3.2-cM region”. Genomics. 66 (1): 119–21. doi:10.1006/geno.2000.6192. PMID10843814.
Schönherr E, Broszat M, Brandan E, Bruckner P, Kresse H (July 1998). “Decorin core protein fragment Leu155-Val260 interacts with TGF-beta but does not compete for decorin binding to type I collagen”. Arch. Biochem. Biophys. 355 (2): 241–8. doi:10.1006/abbi.1998.0720. PMID9675033.
Ebner R, Chen RH, Lawler S, Zioncheck T, Derynck R (November 1993). “Determination of type I receptor specificity by the type II receptors for TGF-beta or activin”. Science. 262 (5135): 900–2. Bibcode:1993Sci…262..900E. doi:10.1126/science.8235612. PMID8235612.
Reinhold D, Wrenger S, Kähne T, Ansorge S (1999). “HIV-1 Tat: immunosuppression via TGF-beta1 induction”. Immunol. Today. 20 (8): 384–5. doi:10.1016/S0167-5699(99)01497-8. PMID10431160.
Yamada Y (2001). “Association of polymorphisms of the transforming growth factor-beta1 gene with genetic susceptibility to osteoporosis”. Pharmacogenetics. 11 (9): 765–71. doi:10.1097/00008571-200112000-00004. PMID11740340.
Chen W, Wahl SM (2002). “TGF-β: Receptors, Signaling Pathways and Autoimmunity”. TGF-beta: receptors, signaling pathways and autoimmunity. Curr. Dir. Autoimmun. Current Directions in Autoimmunity. Vol. 5. pp. 62–91. doi:10.1159/000060548. ISBN978-3-8055-7308-5. PMID11826761.
Marone M, Bonanno G, Rutella S, Leone G, Scambia G, Pierelli L (2002). “Survival and cell cycle control in early hematopoiesis: role of bcl-2, and the cyclin dependent kinase inhibitors P27 and P21”. Leuk. Lymphoma. 43 (1): 51–7. doi:10.1080/10428190210195. PMID11908736. S2CID28490341.
McGowan TA, Zhu Y, Sharma K (2004). “Transforming growth factor-beta: a clinical target for the treatment of diabetic nephropathy”. Curr. Diab. Rep. 4 (6): 447–54. doi:10.1007/s11892-004-0055-z. PMID15539010. S2CID45122439.
TGF beta 2 – TGFB2 – Transforming growth factor-beta 2 (TGF-β2) is a secreted protein known as a cytokine that performs many cellular functions and has a vital role during embryonic development (alternative names: Glioblastoma-derived T-cell suppressor factor, G-TSF, BSC-1 cell growth inhibitor, Polyergin, Cetermin). It is an extracellularglycosylated protein. It is known to suppress the effects of interleukin dependent T-celltumors. There are two named isoforms of this protein, created by alternative splicing of the same gene (i.e., TGFB2).
Noma T, Glick AB, Geiser AG, et al. (1992). “Molecular cloning and structure of the human transforming growth factor-beta 2 gene promoter”. Growth Factors. 4 (4): 247–55. doi:10.3109/08977199109043910. PMID1764261.
Bodmer S, Podlisny MB, Selkoe DJ, et al. (1990). “Transforming growth factor-beta bound to soluble derivatives of the beta amyloid precursor protein of Alzheimer’s disease”. Biochem. Biophys. Res. Commun. 171 (2): 890–7. doi:10.1016/0006-291X(90)91229-L. PMID2119582.
Webb NR, Madisen L, Rose TM, Purchio AF (1989). “Structural and sequence analysis of TGF-beta 2 cDNA clones predicts two different precursor proteins produced by alternative mRNA splicing”. DNA. 7 (7): 493–7. doi:10.1089/dna.1.1988.7.493. PMID2850146.
Madisen L, Webb NR, Rose TM, et al. (1988). “Transforming growth factor-beta 2: cDNA cloning and sequence analysis”. DNA. 7 (1): 1–8. doi:10.1089/dna.1988.7.1. PMID3162414.
Barton DE, Foellmer BE, Du J, et al. (1989). “Chromosomal mapping of genes for transforming growth factors beta 2 and beta 3 in man and mouse: dispersion of TGF-beta gene family”. Oncogene Res. 3 (4): 323–31. PMID3226728.
Philip A, Bostedt L, Stigbrand T, O’Connor-McCourt MD (1994). “Binding of transforming growth factor-beta (TGF-beta) to pregnancy zone protein (PZP). Comparison to the TGF-beta-alpha 2-macroglobulin interaction”. Eur. J. Biochem. 221 (2): 687–93. doi:10.1111/j.1432-1033.1994.tb18781.x. PMID7513640.
Herpin A, Lelong C, Favrel P (May 2004). “Transforming growth factor-beta-related proteins: an ancestral and widespread superfamily of cytokines in metazoans”. Developmental and Comparative Immunology. 28 (5): 461–85. doi:10.1016/j.dci.2003.09.007. PMID15062644.
Taya Y, O’Kane S, Ferguson MW (Sep 1999). “Pathogenesis of cleft palate in TGF-beta3 knockout mice”. Development. 126 (17): 3869–79. doi:10.1242/dev.126.17.3869. PMID10433915.
Kaartinen V, Voncken JW, Shuler C, Warburton D, Bu D, Heisterkamp N, Groffen J (Dec 1995). “Abnormal lung development and cleft palate in mice lacking TGF-beta 3 indicates defects of epithelial-mesenchymal interaction”. Nature Genetics. 11 (4): 415–21. doi:10.1038/ng1295-415. PMID7493022. S2CID22365206.
De Crescenzo G, Pham PL, Durocher Y, O’Connor-McCourt MD (May 2003). “Transforming growth factor-beta (TGF-beta) binding to the extracellular domain of the type II TGF-beta receptor: receptor capture on a biosensor surface using a new coiled-coil capture system demonstrates that avidity contributes significantly to high affinity binding”. Journal of Molecular Biology. 328 (5): 1173–83. doi:10.1016/S0022-2836(03)00360-7. PMID12729750.
Hart PJ, Deep S, Taylor AB, Shu Z, Hinck CS, Hinck AP (Mar 2002). “Crystal structure of the human TbetaR2 ectodomain–TGF-beta3 complex”. Nature Structural Biology. 9 (3): 203–8. doi:10.1038/nsb766. PMID11850637. S2CID13322593.
Barton DE, Foellmer BE, Du J, Tamm J, Derynck R, Francke U (1989). “Chromosomal mapping of genes for transforming growth factors beta 2 and beta 3 in man and mouse: dispersion of TGF-beta gene family”. Oncogene Research. 3 (4): 323–31. PMID3226728.
Kaartinen V, Voncken JW, Shuler C, Warburton D, Bu D, Heisterkamp N, Groffen J (Dec 1995). “Abnormal lung development and cleft palate in mice lacking TGF-beta 3 indicates defects of epithelial-mesenchymal interaction”. Nature Genetics. 11 (4): 415–21. doi:10.1038/ng1295-415. PMID7493022. S2CID22365206.
Nishida K, Sotozono C, Adachi W, Yamamoto S, Yokoi N, Kinoshita S (Mar 1995). “Transforming growth factor-beta 1, -beta 2 and -beta 3 mRNA expression in human cornea”. Current Eye Research. 14 (3): 235–41. doi:10.3109/02713689509033520. PMID7796607.
Rampazzo A, Nava A, Danieli GA, Buja G, Daliento L, Fasoli G, Scognamiglio R, Corrado D, Thiene G (Jun 1994). “The gene for arrhythmogenic right ventricular cardiomyopathy maps to chromosome 14q23-q24”. Human Molecular Genetics. 3 (6): 959–62. doi:10.1093/hmg/3.6.959. PMID7951245.
Zhao Y, Chegini N, Flanders KC (Oct 1994). “Human fallopian tube expresses transforming growth factor (TGF beta) isoforms, TGF beta type I-III receptor messenger ribonucleic acid and protein, and contains [125I]TGF beta-binding sites”. The Journal of Clinical Endocrinology and Metabolism. 79 (4): 1177–84. doi:10.1210/jcem.79.4.7962292. PMID7962292.
Ambros RA, Kallakury BV, Malfetano JH, Mihm MC (Oct 1996). “Cytokine, cell adhesion receptor, and tumor suppressor gene expression in vulvar squamous carcinoma: correlation with prominent fibromyxoid stromal response”. International Journal of Gynecological Pathology. 15 (4): 320–5. doi:10.1097/00004347-199610000-00004. PMID8886879.
Djonov V, Ball RK, Graf S, Mottaz AE, Arnold AM, Flanders K, Studer UE, Merz VW (May 1997). “Transforming growth factor-beta 3 is expressed in nondividing basal epithelial cells in normal human prostate and benign prostatic hyperplasia, and is no longer detectable in prostate carcinoma”. The Prostate. 31 (2): 103–9. doi:10.1002/(SICI)1097-0045(19970501)31:2<103::AID-PROS5>3.0.CO;2-O. PMID9140123. S2CID22734222.
Mori T, Kawara S, Shinozaki M, Hayashi N, Kakinuma T, Igarashi A, Takigawa M, Nakanishi T, Takehara K (Oct 1999). “Role and interaction of connective tissue growth factor with transforming growth factor-beta in persistent fibrosis: A mouse fibrosis model”. Journal of Cellular Physiology. 181 (1): 153–9. doi:10.1002/(SICI)1097-4652(199910)181:1<153::AID-JCP16>3.0.CO;2-K. PMID10457363. S2CID21284888.
A fourth member, TGF beta 4, has been identified in birds – TGRB4 (synonyms: endometrial bleeding associated factor beta-4 (EBAF)[citation needed], Lefty preproprotein[citation needed], LEFTA[citation needed]; Left-Right Determination Factor 2; LEFTYA; Left-Right Determination Factor A; Transforming Growth Factor Beta-4; Protein Lefty-2; Protein Lefty-A).
Roberts, Anita B.; Kim, Seong-Jin; Noma, Takafumi; Glick, Adam B.; Lafyatis, Robert; Lechleider, Robert; Jakowlew, Sonia B.; Geiser, Andrew; O’Reilly, Michael A.; Danielpour, David; Sporn, Michael B. (2007). “Multiple Forms of TGF-β: Distinct Promoters and Differential Expression”. Ciba Foundation Symposium 157 – Clinical Applications of TGF-β. Novartis Foundation Symposia. Vol. 157. pp. 7–28. doi:10.1002/9780470514061.ch2. ISBN978-0-470-51406-1. PMID1906395.
Tabibzadeh S, Kothapalli R, Buyuksal I (July 1997). “Distinct tumor specific expression of TGFB4 (ebaf)*, a novel human gene of the TGF-beta superfamily”. Frontiers in Bioscience. 2: a18-25. doi:10.2741/a158. PMID9230066.
A fifth member of the subfamily, TGFB5, has been identified only in frogs.
Roberts, Anita B.; Kim, Seong-Jin; Noma, Takafumi; Glick, Adam B.; Lafyatis, Robert; Lechleider, Robert; Jakowlew, Sonia B.; Geiser, Andrew; O’Reilly, Michael A.; Danielpour, David; Sporn, Michael B. (2007). “Multiple Forms of TGF-β: Distinct Promoters and Differential Expression”. Ciba Foundation Symposium 157 – Clinical Applications of TGF-β. Novartis Foundation Symposia. Vol. 157. pp. 7–28. doi:10.1002/9780470514061.ch2. ISBN978-0-470-51406-1. PMID1906395.
The peptide structures of the TGF-β isoforms are highly similar (homologies on the order of 70–80%). They are all encoded as large protein precursors; TGF-β1 contains 390 amino acids and TGF-β2 and TGF-β3 each contain 412 amino acids. They each have an N-terminal signal peptide of 20–30 amino acids that they require for secretion from a cell, a pro-region called latency-associated peptide (LAP – Alias: Pro-TGF beta 1, LAP/TGF beta 1), and a 112-114 amino acid C-terminal region that becomes the mature TGF-β molecule following its release from the pro-region by proteolytic cleavage. The mature TGF-β protein dimerizes to produce a 25 KDa active protein with many conserved structural motifs. TGF-β has nine cysteine residues that are conserved among its family. Eight form disulfide bonds within the protein to create a cysteine knot structure characteristic of the TGF-β superfamily. The ninth cysteine forms a disulfide bond with the ninth cysteine of another TGF-β protein to produce a dimer. Many other conserved residues in TGF-β are thought to form secondary structure through hydrophobic interactions. The region between the fifth and sixth conserved cysteines houses the most divergent area of TGF-β proteins that is exposed at the surface of the protein and is implicated in receptor binding and specificity of TGF-β.
Herpin A, Lelong C, Favrel P (May 2004). “Transforming growth factor-beta-related proteins: an ancestral and widespread superfamily of cytokines in metazoans”. Developmental and Comparative Immunology. 28 (5): 461–85. doi:10.1016/j.dci.2003.09.007. PMID15062644.
Although TGF-β is important in regulating crucial cellular activities, only a few TGF-β activating pathways are currently known, and the full mechanism behind the suggested activation pathways is not yet well understood. Some of the known activating pathways are cell or tissue specific, while some are seen in multiple cell types and tissues. Proteases, integrins, pH, and reactive oxygen species are just few of the currently known factors that can activate TGF-β, as discussed below. It is well known that perturbations of these activating factors can lead to unregulated TGF-β signaling levels that may cause several complications including inflammation, autoimmune disorders, fibrosis, cancer and cataracts. In most cases, an activated TGF-β ligand will initiate the TGF-β signaling cascade as long as TGF-β receptors I and II are available for binding. This is due to a high affinity between TGF-β and its receptors, suggesting why the TGF-β signaling recruits a latency system to mediate its signaling.
Wipff PJ, Hinz B (September 2008). “Integrins and the activation of latent transforming growth factor beta1 – an intimate relationship”. European Journal of Cell Biology. 87 (8–9): 601–15. doi:10.1016/j.ejcb.2008.01.012. PMID18342983.
Plasmin and a number of matrix metalloproteinases (MMP) play a key role in promoting tumor invasion and tissue remodeling by inducing proteolysis of several ECM components. The TGF-β activation process involves the release of the LLC from the matrix, followed by further proteolysis of the LAP to release TGF-β to its receptors. MMP-9 and MMP-2 are known to cleave latent TGF-β. The LAP complex contains a protease-sensitive hinge region which can be the potential target for this liberation of TGF-β. Despite the fact that MMPs have been proven to play a key role in activating TGF-β, mice with mutations in MMP-9 and MMP-2 genes can still activate TGF-β and do not show any TGF-β deficiency phenotypes, this may reflect redundancy among the activating enzymes suggesting that other unknown proteases might be involved.
Acidic conditions can denature the LAP. Treatment of the medium with extremes of pH (1.5 or 12) resulted in significant activation of TGF-β as shown by radio-receptor assays, while mild acid treatment (pH 4.5) yielded only 20-30% of the activation achieved by pH 1.5.
The structure of LAP is important in maintaining its function. Structure modification of LAP can lead to disturb the interaction between LAP and TGF-β and thus activating it. Factors that may cause such modification may include hydroxyl radicals from reactive oxygen species (ROS). TGF-β was rapidly activated after in vivo radiation exposure ROS.
Thrombospondin-1 (TSP-1) is a matricellular glycoprotein found in plasma of healthy patients with levels in the range of 50–250 ng/ml. TSP-1 levels are known to increase in response to injury and during development. TSP-1 activates latent TGF-beta by forming direct interactions with the latent TGF-β complex and induces a conformational rearrangement preventing it from binding to the matured TGF-β.
Booth WJ, Berndt MC (July 1987). “Thrombospondin in clinical disease states”. Seminars in Thrombosis and Hemostasis. 13 (3): 298–306. doi:10.1055/s-2007-1003505. PMID3317840.
Raugi GJ, Olerud JE, Gown AM (December 1987). “Thrombospondin in early human wound tissue”. The Journal of Investigative Dermatology. 89 (6): 551–4. doi:10.1111/1523-1747.ep12461198. PMID3680981.
Murphy-Ullrich JE, Poczatek M (2000). “Activation of latent TGF-beta by thrombospondin-1: mechanisms and physiology”. Cytokine & Growth Factor Reviews. 11 (1–2): 59–69. doi:10.1016/S1359-6101(99)00029-5. PMID10708953.
Activation by Alpha(V) containing integrins
The general theme of integrins participating in latent TGF-β1 activation arose from studies that examined mutations/knockouts of β6 integrin, αV integrin, β8 integrin and in LAP. These mutations produced phenotypes that were similar to phenotypes seen in TGF-β1 knockout mice. Currently there are two proposed models of how αV containing integrins can activate latent TGF-β1; the first proposed model is by inducing conformational change to the latent TGF-β1 complex and hence releasing the active TGF-β1 and the second model is by a protease-dependent mechanism.
Wipff PJ, Hinz B (September 2008). “Integrins and the activation of latent transforming growth factor beta1 – an intimate relationship”. European Journal of Cell Biology. 87 (8–9): 601–15. doi:10.1016/j.ejcb.2008.01.012. PMID18342983.
αVβ6 integrin was the first integrin to be identified as TGF-β1 activator. LAPs contain an RGD motif which is recognized by vast majority of αV containing integrins, and αVβ6 integrin can activate TGF-β1 by binding to the RGD motif present in LAP-β1 and LAP-β3. Upon binding, it induces adhesion-mediated cell forces that are translated into biochemical signals which can lead to liberation/activation of TGFb from its latent complex. This pathway has been demonstrated for activation of TGF-β in epithelial cells and does not associate MMPs.
Because MMP-2 and MMP-9 can activate TGF-β through proteolytic degradation of the latent TGF beta complex, αV containing integrins activate TGF-β1 by creating a close connection between the latent TGF-β complex and MMPs. Integrins αVβ6 and αVβ3 are suggested to simultaneously bind the latent TGF-β1 complex and proteinases, simultaneous inducing conformational changes of the LAP and sequestering proteases to close proximity. Regardless of involving MMPs, this mechanism still necessitate the association of integrins and that makes it a non proteolytic pathway.
Wipff PJ, Hinz B (September 2008). “Integrins and the activation of latent transforming growth factor beta1 – an intimate relationship”. European Journal of Cell Biology. 87 (8–9): 601–15. doi:10.1016/j.ejcb.2008.01.012. PMID18342983.
Perlman R, Schiemann WP, Brooks MW, Lodish HF, Weinberg RA (August 2001). “TGF-beta-induced apoptosis is mediated by the adapter protein Daxx that facilitates JNK activation”. Nature Cell Biology. 3 (8): 708–14. doi:10.1038/35087019. PMID11483955. S2CID20435808.
Hofmann TG, Stollberg N, Schmitz ML, Will H (December 2003). “HIPK2 regulates transforming growth factor-beta-induced c-Jun NH(2)-terminal kinase activation and apoptosis in human hepatoma cells”. Cancer Research. 63 (23): 8271–7. PMID14678985.
Inman, Gareth J.; Nicolás, Francisco J.; Callahan, James F.; Harling, John D.; Gaster, Laramie M.; Reith, Alastair D.; Laping, Nicholas J.; Hill, Caroline S. (1 July 2002). “SB-431542 Is a Potent and Specific Inhibitor of Transforming Growth Factor-β Superfamily Type I Activin Receptor-Like Kinase (ALK) Receptors ALK4, ALK5, and ALK7”. Molecular Pharmacology. 62 (1): 65–74. doi:10.1124/mol.62.1.65. PMID12065756. S2CID15185199.
Mukundan, Ananya; Byeon, Chang-Hyeock; Hinck, Cynthia S.; Smyth, Danielle J.; Maizels, Rick M.; Hinck, Andrew P. (2020-12-09). “Structure-based mapping of the TβRI and TβRII receptor binding sites of the parasitic TGF-β mimic, Hp-TGM”. doi:10.1101/2020.12.08.416701. S2CID229293860.
Morishima N, Mizoguchi I, Takeda K, Mizuguchi J, Yoshimoto T (August 2009). “TGF-beta is necessary for induction of IL-23R and Th17 differentiation by IL-6 and IL-23”. Biochemical and Biophysical Research Communications. 386 (1): 105–10. doi:10.1016/j.bbrc.2009.05.140. PMID19501566.
Blobe GC, Schiemann WP, Lodish HF (May 2000). “Role of transforming growth factor beta in human disease”. The New England Journal of Medicine. 342 (18): 1350–8. doi:10.1056/NEJM200005043421807. PMID10793168.
Bone morphogenetic proteins (BMPs) are a group of growth factors also known as cytokines and as metabologens. Originally discovered by their ability to induce the formation of bone and cartilage, BMPs are now considered to constitute a group of pivotal morphogenetic signals, orchestrating tissue architecture throughout the body. The important functioning of BMP signals in physiology is emphasized by the multitude of roles for dysregulated BMP signaling in pathological processes. Cancerous disease often involves misregulation of the BMP signaling system. Absence of BMP signaling is, for instance, an important factor in the progression of colon cancer, and conversely, overactivation of BMP signaling following reflux-induced esophagitis provokes Barrett’s esophagus and is thus instrumental in the development of esophageal adenocarcinoma.
Reddi AH, Reddi A (2009). “Bone morphogenetic proteins (BMPs): from morphogens to metabologens”. Cytokine & Growth Factor Reviews. 20 (5–6): 341–2. doi:10.1016/j.cytogfr.2009.10.015. PMID19900831.
Milano F, van Baal JW, Buttar NS, Rygiel AM, de Kort F, DeMars CJ, Rosmolen WD, Bergman JJ, VAn Marle J, Wang KK, Peppelenbosch MP, Krishnadath KK (Jun 2007). “Bone morphogenetic protein 4 expressed in esophagitis induces a columnar phenotype in esophageal squamous cells”. Gastroenterology. 132 (7): 2412–21. doi:10.1053/j.gastro.2007.03.026. PMID17570215.
Signal transduction through BMPRs results in mobilization of members of the SMAD family of proteins. The signaling pathways involving BMPs, BMPRs and SMADs are important in the development of the heart, central nervous system, and cartilage, as well as post-natal bone development.
They have an important role during embryonic development on the embryonic patterning and early skeletal formation. As such, disruption of BMP signaling can affect the body plan of the developing embryo. For example, BMP4 and its inhibitors noggin and chordin help regulate polarity of the embryo (i.e. back to front patterning). Specifically BMP-4 and its inhibitors play a major role in neurulation and the development of the neural plate. BMP-4 signals ectoderm cells to develop into skin cells, but the secretion of inhibitors by the underlying mesoderm blocks the action of BMP-4 to allow the ectoderm to continue on its normal course of neural cell development. Additionally, secretion of BMPs by the roof plate in the developing spinal cord helps to specify dorsal sensory interneurons.
As a member of the transforming growth factor-beta superfamily, BMP signaling regulates a variety of embryonic patterning during fetal and embryonic development. For example, BMP signaling controls the early formation of the Mullerian duct (MD) which is a tubular structure in early embryonic developmental stage and eventually becomes female reproductive tracts. Chemical inhibiting BMP signals in chicken embryo caused a disruption of MD invagination and blocked the epithelial thickening of the MD-forming region, indicating that the BMP signals play a role in early MD development. Moreover, BMP signaling is involved in the formation of foregut and hindgut, intestinal villus patterning, and endocardial differentiation. Villi contribute to increase the effective absorption of nutrients by extending the surface area in small intestine. Gain or lose function of BMP signaling altered the patterning of clusters and emergence of villi in mouse intestinal model. BMP signal derived from myocardium is also involved in endocardial differentiation during heart development. Inhibited BMP signal in zebrafish embryonic model caused strong reduction of endocardial differentiation, but only had little effect in myocardial development. In addition, Notch-Wnt-Bmp crosstalk is required for radial patterning during mouse cochlea development via antagonizing manner.
Mutations in BMPs and their inhibitors are associated with a number of human disorders which affect the skeleton.
BMPs are also involved in adipogenesis and functional regulation of adipose tissue. BMP4 favors white adipogenesis, whereas BMP7 activates brown fat functionality; BMP inhibitors are also involved in this regulation.
Originally, seven such proteins were discovered. Of these, six (BMP2 through BMP7) belong to the Transforming growth factor beta superfamily of proteins. BMP1 is a metalloprotease. Since then, thirteen more BMPs, all of which are in the TGF-beta family, have been discovered, bringing the total to twenty.
Even J, Eskander M, Kang J (Sep 2012). “Bone morphogenetic protein in spine surgery: current and future uses”. The Journal of the American Academy of Orthopaedic Surgeons. 20 (9): 547–52. doi:10.5435/JAAOS-20-09-547. PMID22941797.
The current nomenclature only recognizes 13, as many others are put under the growth differentiation factor naming instead.
*BMP1 does not belong to the TGF-β family of proteins. It is a metalloprotease that acts on procollagen I, II, and III. It is involved in cartilage development.
Acts as a disulfide-linked homodimer and induces bone and cartilage formation. It is a candidate as a retinoid mediator. Plays a key role in osteoblast differentiation.
Regulates the formation of teeth, limbs and bone from mesoderm. It also plays a role in fracture repair, epidermis formation, dorsal-ventral axis formation, and ovarian follical development.
Several BMPs are also named ‘cartilage-derived morphogenetic proteins’ (CDMPs), while others are referred to as ‘growth differentiation factors‘ (GDFs).
Herpin A, Lelong C, Favrel P (2004). “Transforming growth factor-beta-related proteins: an ancestral and widespread superfamily of cytokines in metazoans”. Dev Comp Immunol. 28 (5): 461–85. doi:10.1016/j.dci.2003.09.007. PMID15062644.
Several members of this subfamily have been described, and named GDF1 through GDF15.
Rankin C, Bunton T, Lawler A, Lee S (2000). “Regulation of left-right patterning in mice by growth/differentiation factor-1”. Nat Genet. 24 (3): 262–5. doi:10.1038/73472. PMID10700179. S2CID6787053.
GDF2 (also known as BMP9) induces and maintains the response embryonic basal forebrain cholinergic neurons (BFCN) have to a neurotransmitter called acetylcholine, and regulates iron metabolism by increasing levels of a protein called hepcidin.
O’Keeffe G, Dockery P, Sullivan A (2004). “Effects of growth/differentiation factor 5 on the survival and morphology of embryonic rat midbrain dopaminergic neurones in vitro”. J Neurocytol. 33 (5): 479–88. doi:10.1007/s11068-004-0511-y. PMID15906156. S2CID25940876.
Buxton P, Edwards C, Archer C, Francis-West P (2001). “Growth/differentiation factor-5 (GDF-5) and skeletal development”. J Bone Joint Surg Am. 83-A Suppl 1 (Pt 1): S23–30. PMID11263662.
Francis-West P, Parish J, Lee K, Archer C (1999). “BMP/GDF-signalling interactions during synovial joint development”. Cell Tissue Res. 296 (1): 111–9. doi:10.1007/s004410051272. PMID10199971. S2CID21942870.
Chang C, Hemmati-Brivanlou A (1999). “Xenopus GDF6, a new antagonist of noggin and a partner of BMPs”. Development. 126 (15): 3347–57. doi:10.1242/dev.126.15.3347. PMID10393114.
GDF8 is now officially known as myostatin and controls the growth of muscle tissue.[14]
McPherron AC, Lawler AM, Lee SJ (May 1997). “Regulation of skeletal muscle mass in mice by a new TGF-beta superfamily member”. Nature. 387 (6628): 83–90. doi:10.1038/387083a0. PMID9139826. S2CID4271945.
GDF9, like GDF3, lacks one cysteine relative to other members of the TGF-β superfamily. Its gene expression is limited to the ovaries, and it has a role in ovulation.
Juengel J, Bodensteiner K, Heath D, Hudson N, Moeller C, Smith P, Galloway S, Davis G, Sawyer H, McNatty K (2004). “Physiology of GDF9 and BMP15 signalling molecules”. Anim Reprod Sci. 82–83: 447–60. doi:10.1016/j.anireprosci.2004.04.021. PMID15271472.
GDF10 is closely related to BMP3 and has a roles in head formation and, it is presumed, in skeletal morphogenesis. It is also known as BMP-3b.
Hino J, Kangawa K, Matsuo H, Nohno T, Nishimatsu S (2004). “Bone morphogenetic protein-3 family members and their biological functions”. Front Biosci. 9 (1–3): 1520–9. doi:10.2741/1355. PMID14977563.
Cunningham N, Jenkins N, Gilbert D, Copeland N, Reddi A, Lee S (1995). “Growth/differentiation factor-10: a new member of the transforming growth factor-beta superfamily related to bone morphogenetic protein-3”. Growth Factors. 12 (2): 99–109. doi:10.3109/08977199509028956. PMID8679252.
GDF15 (also known as TGF-PL, MIC-1, PDF, PLAB, and PTGFB) has a role in regulating inflammatory and apoptotic pathways during tissue injury and certain disease processes.
Zimmers T, Jin X, Hsiao E, McGrath S, Esquela A, Koniaris L (2005). “Growth differentiation factor-15/macrophage inhibitory cytokine-1 induction after kidney and lung injury”. Shock. 23 (6): 543–8. PMID15897808.
Activin and inhibin are two closely related proteincomplexes that have almost directly opposite biological effects. Identified in 1986, activin enhances FSHbiosynthesis and secretion, and participates in the regulation of the menstrual cycle.
Vale W, Rivier J, Vaughan J, McClintock R, Corrigan A, Woo W, Karr D, Spiess J (1986). “Purification and characterization of an FSH releasing protein from porcine ovarian follicular fluid”. Nature. 321 (6072): 776–9. Bibcode:1986Natur.321..776V. doi:10.1038/321776a0. PMID3012369. S2CID4365045.
Ling N, Ying SY, Ueno N, Shimasaki S, Esch F, Hotta M, Guillemin R (1986). “Pituitary FSH is released by a heterodimer of the beta-subunits from the two forms of inhibin”. Nature. 321 (6072): 779–82. Bibcode:1986Natur.321..779L. doi:10.1038/321779a0. PMID3086749. S2CID38100413.
Sulyok S, Wankell M, Alzheimer C, Werner S (October 2004). “Activin: an important regulator of wound repair, fibrosis, and neuroprotection”. Molecular and Cellular Endocrinology. 225 (1–2): 127–32. doi:10.1016/j.mce.2004.07.011. PMID15451577. S2CID6943949.
Conversely, inhibin downregulates FSH synthesis and inhibits FSH secretion.
The existence of inhibin was hypothesized as early as 1916; however, it was not demonstrated to exist until Neena Schwartz and Cornelia Channing‘s work in the mid-1970s, after which both proteins were molecularly characterized ten years later.
Activin is a dimer composed of two identical or very similar beta subunits. Inhibin is also a dimer wherein the first component is a beta subunit similar or identical to the beta subunit in activin. However, in contrast to activin, the second component of the inhibin dimer is a more distantly-related alpha subunit. Activin, inhibin and a number of other structurally related proteins such as anti-Müllerian hormone, bone morphogenetic protein, and growth differentiation factor belong to the TGF-β protein superfamily.
Burger HG, Igarashi M (April 1988). “Inhibin: definition and nomenclature, including related substances”. The Journal of Clinical Endocrinology and Metabolism. 66 (4): 885–6. PMID3346366.
The activin and inhibin protein complexes are both dimeric in structure, and, in each complex, the two monomers are linked to one another by a single disulfide bond.
Ying SY (December 1987). “Inhibins and activins: chemical properties and biological activity”. Proceedings of the Society for Experimental Biology and Medicine. 186 (3): 253–64. doi:10.3181/00379727-186-42611a. PMID3122219. S2CID36872324.
In addition, both complexes are derived from the same family of related genes and proteins but differ in their subunit composition.
Burger HG, Igarashi M (April 1988). “Inhibin: definition and nomenclature, including related substances”. The Journal of Clinical Endocrinology and Metabolism. 66 (4): 885–6. PMID3346366.
The alpha and beta subunits share approximately 25% sequence similarity, whereas the similarity between beta subunits is approximately 65%.
In mammals, four beta subunits have been described, called activin βA, activin βB, activin βC and activin βE. Activin βA and βB are identical to the two beta subunits of inhibin. A fifth subunit, activin βD, has been described in Xenopus laevis. Two activin βA subunits give rise to activin A, one βA, and one βB subunit gives rise to activin AB, and so on. Various, but not all theoretically possible, heterodimers have been described.
Activin is strongly expressed in wounded skin, and overexpression of activin in epidermis of transgenicmice improves wound healing and enhances scar formation. Its action in wound repair and skin morphogenesis is through stimulation of keratinocytes and stromal cells in a dose-dependent manner.
Activin also regulates the morphogenesis of branching organs such as the prostate, lung, and especially kidney. Activin A increased the expression level of type-I collagen suggesting that activin A acts as a potent activator of fibroblasts.
Lack of activin during development results in neural developmental defects.
Upregulation of Activin A drives pluripotent stem cells into a mesoendodermal fate, and thus provides a useful tool for stem cell differentiation and organoid formation.
As with other members of the superfamily, activins interact with two types of cell surface transmembrane receptors (Types I and II) which have intrinsic serine/threonine kinase activities in their cytoplasmic domains:
Activin binds to the Type II receptor and initiates a cascade reaction that leads to the recruitment, phosphorylation, and activation of Type I activin receptor. This then interacts with and then phosphorylates SMAD2 and SMAD3, two of the cytoplasmic SMAD proteins.
Smad3 then translocates to the nucleus and interacts with SMAD4 through multimerization, resulting in their modulation as transcription factor complexes responsible for the expression of a large variety of genes.
Inhibin
In contrast to activin, much less is known about the mechanism of action of inhibin, but may involve competing with activin for binding to activin receptors and/or binding to inhibin-specific receptors.
A mutation in the gene for the activin receptor ACVR1 results in fibrodysplasia ossificans progressiva, a fatal disease that causes muscle and soft tissue to gradually be replaced by bone tissue. This condition is characterized by the formation of an extra skeleton that produces immobilization and eventually death by suffocation. The mutation in ACVR1 causes activin A, which normally acts as an antagonist of the receptor and blocks osteogenesis (bone growth), to behave as an agonist of the receptor and to induce hyperactive bone growth.
Shore EM, Xu M, Feldman GJ, Fenstermacher DA, Cho TJ, Choi IH, Connor JM, Delai P, Glaser DL, LeMerrer M, Morhart R, Rogers JG, Smith R, Triffitt JT, Urtizberea JA, Zasloff M, Brown MA, Kaplan FS (May 2006). “A recurrent mutation in the BMP type I receptor ACVR1 causes inherited and sporadic fibrodysplasia ossificans progressiva”. Nature Genetics. 38 (5): 525–527. doi:10.1038/ng1783. PMID16642017. S2CID41579747.
Quantification of inhibin A is part of the prenatal quad screen that can be administered during pregnancy at a gestational age of 16–18 weeks. An elevated inhibin A (along with an increased beta-hCG, decreased AFP, and a decreased estriol) is suggestive of the presence of a fetus with Down syndrome.
Aitken DA, Wallace EM, Crossley JA, Swanston IA, van Pareren Y, van Maarle M, Groome NP, Macri JN, Connor JM (May 1996). “Dimeric inhibin A as a marker for Down’s syndrome in early pregnancy”. The New England Journal of Medicine. 334 (19): 1231–6. doi:10.1056/NEJM199605093341904. PMID8606718.
As a screening test, abnormal quad screen test results need to be followed up with more definitive tests.
Robertson DM, Pruysers E, Jobling T (April 2007). “Inhibin as a diagnostic marker for ovarian cancer”. Cancer Letters. 249 (1): 14–7. doi:10.1016/j.canlet.2006.12.017. PMID17320281.
Robertson DM, Pruysers E, Burger HG, Jobling T, McNeilage J, Healy D (October 2004). “Inhibins and ovarian cancer”. Molecular and Cellular Endocrinology. 225 (1–2): 65–71. doi:10.1016/j.mce.2004.02.014. PMID15451569. S2CID33801243
Inhibin B may be used as a marker of spermatogenesis function and male infertility. The mean serum inhibin B level is significantly higher among fertile men (approximately 140 pg/mL) than in infertile men (approximately 80 pg/mL).
In men with azoospermia, a positive test for inhibin B slightly raises the chances for successfully achieving pregnancy through testicular sperm extraction (TESE), although the association is not very substantial, having a sensitivity of 0.65 (95% confidence interval [CI]: 0.56–0.74) and a specificity of 0.83 (CI: 0.64–0.93) for prediction the presence of sperm in the testes in non-obstructive azoospermia.
Lefty (left-right determination factors) are a class of proteins that are closely related members of the TGF-beta superfamily of growth factors. These proteins are secreted and play a role in left-right asymmetry determination of organ systems during development. Mutations of the genes encoding these proteins have been associated with left-right axis malformations, particularly in the heart and lungs.
Hamada H, Meno C, Watanabe D, Saijoh Y (February 2002). “Establishment of vertebrate left-right asymmetry”. Nat. Rev. Genet. 3 (2): 103–13. doi:10.1038/nrg732. PMID11836504. S2CID20557143.
Lefty, a divergent member of the transforming growth factor-β (TGF beta) superfamily of proteins, was originally discovered in the Hamada lab at the Osaka University using deletion screening of cDNA libraries in P19 embryonic carcinoma cells to find clones that did not differentiate when induced to differentiate using retinoic acid. From these screens, researchers found one gene that was a tentative member of the TGF-beta superfamily that was predominantly expressed on the left side the embryo and aptly named it lefty.
Meno C, Saijoh Y, Fujii H, Ikeda M, Yokoyama T, Yokoyama M, Toyoda Y, Hamada H (May 1996). “Left-right asymmetric expression of the TGF beta-family member lefty in mouse embryos”. Nature. 381 (6578): 151–5. Bibcode:1996Natur.381..151M. doi:10.1038/381151a0. PMID8610011. S2CID4345275.
Like other members of the TGF-beta superfamily, lefty is synthesized as a preproprotein, meaning that the protein is proteolytically cleaved and excreted to produce the active form of the protein. However, lefty has only 20-25% sequence similarity with other members of the TGF-beta superfamily. Lefty is conserved in all vertebrates and many species have more than one homologue. Humans and mice, for instance have two homologues, Lefty 1 and Lefty 2, whose differential expression leads to distinct purposes while the mechanism of action is conserved.
Lefty proteins function as an antagonist of the Nodal Signaling pathway. Nodal is another signaling protein which is responsible for gastrulation, left-right patterning and induction of the primitive node. As NODAL protein diffuse through an embryo, it triggers Nodal Signaling within tissues with the required receptors and coreceptors. Activated nodal signaling leads to the transcription of the lefty gene. The protein is then expressed, proteolytically cleaved, and finally secreted. Secreted lefty binds to EGF-CFC proteins like one-eyed pinhead in zebrafish keeping the essential cofactor from associating with NODAL/ Activin-like receptor complex. This will effectually block Nodal Signaling. During induction of the primitive streak, lefty confines Nodal activity to the posterior end of the embryo, establishing a posterior signaling center and inducing the formation of the primitive streak and mesoderm. (See Nodal Signaling or TGF beta signaling pathway for more information on the nodal signaling pathway.)
Carlson, Bruce M. “Formation of Germ Layers and Early Derivatives.” Human Embryology and Developmental Biology. Philadelphia, Pennsylvania: Mosby/Elsevier, 2009. 91-95. Print.
There are many differences between the left and right sides, including heart and lung positioning. Mutations in these genes cause incorrect positioning of these organs (e.g., situs inversus), or in the case of constitutively inactive lefty, the embryo becomes entirely mesoderm and fails to pattern or develop. During vertebrate development, lefty proteins regulate left-right asymmetry by controlling the spatiotemporal influence of the NODAL protein. Lefty1 in the ventral midline prevents the Cerberus (paracrine factor or “Caronte”) signal from passing to the right side of the embryo.
Hamada H, Meno C, Watanabe D, Saijoh Y (February 2002). “Establishment of vertebrate left-right asymmetry”. Nat. Rev. Genet. 3 (2): 103–13. doi:10.1038/nrg732. PMID11836504. S2CID20557143
This spatiotemporal control is achieved by using two sources of excreted lefty. While lefty is produced in response to activated nodal signaling, it is also produced and secreted in the anterior visceral endoderm (AVE). The balance of lefty from the AVE and from Nodal Signaling results in the patterning of the embryo and left-right asymmetry.
Takaoka K, Yamamoto M, Hamada H (August 2007). “Origin of body axes in the mouse embryo”. Curr. Opin. Genet. Dev. 17 (4): 344–50. doi:10.1016/j.gde.2007.06.001. PMID17646095.
Proper functioning of Lefty is crucial to the proper development of the heart, lungs, spleen, and liver. Mutations in Lefty, called Lefty-A, are associated with left-right patterning defects. This mutation may cause congenital heart defects due to malformation, interrupted inferior vena cava, and lack of lung asymmetry (left pulmonary isomerism).
Carlson, Bruce M. “Formation of Germ Layers and Early Derivatives.” Human Embryology and Developmental Biology. Philadelphia, Pennsylvania: Mosby/Elsevier, 2009. 91-95. Print.
Tabibzadeh S (2005). “Role of EBAF/Lefty in implantation and uterine bleeding”. Ernst Schering Res. Found. Workshop. Ernst Schering Research Foundation Workshop. 52 (52): 159–89. doi:10.1007/3-540-27147-3_8. ISBN978-3-540-23089-2. PMID15704472.
Lefty-1 is a regulatory gene that plays a vital role in the determination of the left-right internal asymmetry observed in mammals. The lefty-1 protein works in tandem with two other genes: lefty-2 and nodal. As the primitive node migrates towards the cranial end of the embryo during development, its cilia preferentially sling lefty-2 and nodal towards the left side of the embryo.
Hashimoto M, Shinohara K, Wang J, Ikeuchi S, Yoshiba S, Meno C, Nonaka S, Takada S, Hatta K, Wynshaw-Boris A, Hamada H (February 2010). “Planar polarization of node cells determines the rotational axis of node cilia”. Nature Cell Biology. 12 (2): 170–6. doi:10.1038/ncb2020. PMID20098415. S2CID6379844.
These two genes encode for “leftness”, and initiate the formation of the heart, spleen, and other internal organs that are found on the left side in a typical human being. Lefty-1 protein can be viewed as a barrier between the left and right portions of the embryo that prevents the diffusion of lefty-2 and nodal to the right side. This ensures that the left-determining molecules are confined to their correct developmental domain. A variety of defects were observed in mice that had lefty-1 deleted, including left pulmonary isomerism, situs inversus, and atrial septal defect.
The high incidence of left pulmonary isomerism in the knockout mice indicates that lefty-1 itself is not involved in encoding for leftness, but simply ensures the correct compartmentation of the left-determining molecules. In the absence of the lefty-1 barrier, lefty-2 and nodal are free to diffuse to the right side and initiate the development of a left lung that was meant to be limited to the left side of the thoracic cavity.
Further reading
Carlson BM (2014). “Formation of germ layers and early derivatives.”. Human Embryology and Developmental Biology. Philadelphia, Pennsylvania: Mosby/Elsevier. pp. 75–91. ISBN978-0-323-08279-2.
Sakuma R, Ohnishi Yi Y, Meno C, Fujii H, Juan H, Takeuchi J, Ogura T, Li E, Miyazono K, Hamada H (April 2002). “Inhibition of Nodal signalling by Lefty mediated through interaction with common receptors and efficient diffusion”. Genes to Cells: Devoted to Molecular & Cellular Mechanisms. 7 (4): 401–12. doi:10.1046/j.1365-2443.2002.00528.x. PMID11952836. S2CID19320756.
4. A group encompassing various divergent members?
Transforming growth factor-beta (TGF-beta) is a multifunctional peptide that controls proliferation, differentiation and other functions in many cell types.
TGF-beta-1 is a peptide of 112 amino acid residues derived by proteolytic cleavage from the C-terminal of a precursor protein. These proteins interact with a conserved family of cell surface serine/threonine-specific protein kinase receptors, and generate intracellular signals using a conserved family of proteins called SMADs. They play fundamental roles in the regulation of basic biological processes such as growth, development, tissue homeostasis and regulation of the immune system.
Herpin A, Lelong C, Favrel P (May 2004). “Transforming growth factor-beta-related proteins: an ancestral and widespread superfamily of cytokines in metazoans”. Dev. Comp. Immunol. 28 (5): 461–85. doi:10.1016/j.dci.2003.09.007. PMID15062644.
Structure
Proteins from the TGF-beta superfamily are only active as homo- or heterodimer; the two chains being linked by a single disulfide bond. From X-ray studies of TGF-beta-2, it is known that all the other cysteines are involved in intrachain disulfide bonds.
As shown in the following schematic representation, there are four disulfide bonds in the TGF-beta’s and in inhibin beta chains, while the other members of this superfamily lack the first bond.
Herpin A, Lelong C, Favrel P (May 2004). “Transforming growth factor-beta-related proteins: an ancestral and widespread superfamily of cytokines in metazoans”. Dev. Comp. Immunol. 28 (5): 461–85. doi:10.1016/j.dci.2003.09.007. PMID15062644.
Andres JL, DeFalcis D, et al. (1992). “Binding of two growth factor families to separate domains of the proteoglycan betaglycan”. J. Biol. Chem. 267 (9): 5927–5930. PMID1556106.
Intercellular signaling peptides and proteins / ligands
{kind=link}
Leave a Reply